




%20(3)-Photoroom.png)

Repensando el autismo: el impacto de los factores de riesgo maternos en el desarrollo del autismo
- autismoenvivo

- 9 sept 2024
- 23 Min. de lectura

POR JIANPING LU,1,2 ZICHEN WANG,3 YUJIE LIANG,1,3 Y PAUL YAO1
Fuente: NIH | 15/02/2022
Figuras y tablas: NIH
1 Department of Child and Adolescent Psychiatry, Shenzhen Kangning Hospital, Shenzhen Mental Health Center, Shenzhen Key Laboratory for Psychological Healthcare and Shenzhen Institute of Mental Health, Shenzhen, Guangdong, China
2 Faculty of Mental Health, Shenzhen University, Shenzhen, Guangdong, China
3 Department of Biomedical Engineering, Southern University of Science and Technology, Shenzhen 518055, China
Address correspondence to: Yujie Liang and Paul Yao, Department of Child and Adolescent Psychiatry, Shenzhen Kangning Hospital, Shenzhen Mental Health Center, Shenzhen Key Laboratory for Psychological Healthcare and Shenzhen Institute of Mental Health, No. 1080 Cuizhu Road, Luohu District, Shenzhen, Guangdong, China. Tel: +86-13267085280; E-mail: moc.621@eijygnail (YJL); Tel: +86-0755-258374490; E-mail: moc.oohay@211silisav (PY)
Los factores ambientales, que incluyen circunstancias o acontecimientos prenatales, desempeñan todos ellos un papel muy importante en el desarrollo temprano del autismo, aunque el mecanismo exacto sigue estando en gran medida sin determinar.
Resumen
Los trastornos del espectro autista (TEA) son un grupo de trastornos del neurodesarrollo que duran toda la vida y se caracterizan por déficits cognitivos y alteraciones del desarrollo social y comunicativo, cuya prevalencia ha ido en aumento en las últimas décadas. Estos trastornos pueden ir acompañados de problemas de salud discapacitantes y, a menudo, suponen una carga económica considerable. Las causas y los mecanismos de los TEA aún no se han dilucidado por completo, aunque se ha informado de que los antecedentes genéticos, la modificación epigenética y los factores de riesgo ambientales contribuyen al desarrollo de los TEA.
Los factores ambientales, que incluyen circunstancias o acontecimientos prenatales, desempeñan todos ellos un papel muy importante en el desarrollo temprano del autismo, aunque el mecanismo exacto sigue estando en gran medida sin determinar.
En esta revisión, promovemos un “replanteamiento” del autismo como una enfermedad del neurodesarrollo que se origina en el desarrollo temprano de la vida. Nos centramos en el impacto de los factores de riesgo prenatales y maternos, como la diabetes materna, la exposición prenatal a sustancias químicas y los desequilibrios hormonales durante el embarazo, en el riesgo de desarrollo del TEA en niños y descendientes, identificando importantes bases patológicas y medidas de prevención para décadas futuras.
Nuevas investigaciones centradas en la comprensión del papel de los factores ambientales en la etiología de los TEA impulsarán estrategias innovadoras de intervención y prevención de los factores de riesgo maternos del autismo.
Palabras clave: autismo, factores de riesgo, exposición prenatal.
Introducción
Antes del siglo XXI, los trastornos y enfermedades mentales graves, la lepra y la tuberculosis eran algunos de los principales problemas de salud pública a los que se enfrentaba el mundo. Hoy en día, sólo los trastornos mentales, en particular los trastornos del espectro autista (TEA), siguen teniendo una gran prevalencia mundial y siguen siendo incurables. En los últimos 50 años, la tasa de prevalencia del autismo se ha multiplicado por más de 100, pasando de 1:10000 a 1:59 [1,2]. Sin embargo, a pesar de décadas de investigación, sigue sin haber una cura eficaz o una estrategia de prevención para el autismo.
Por lo tanto, es urgente desarrollar plenamente aplicaciones básicas y clínicas para la investigación del autismo que rompan con el pensamiento tradicional para integrar conceptualizaciones nuevas e innovadoras de la etiología de los TEA.
Las investigaciones anteriores sobre el autismo se han centrado generalmente en la identificación de factores genéticos relacionados con el autismo, pero amplios estudios realizados en los últimos años han descubierto que los factores ambientales también están implicados en gran medida. Sobre una base genética relativamente heterogénea, la red epigenética construida por la interacción de factores ambientales y genéticos es un eslabón clave para determinar el riesgo de desarrollo del TEA.
Estudios familiares, análisis de pedigrí y estudios de gemelos han sugerido previamente que el autismo tiene una fuerte base genética, siendo los factores genéticos responsables del 30-40% de los casos de autismo [3,4]. Sin embargo, es importante reconocer que la causa principal no es únicamente genética, sino que la epigenética puede desempeñar un papel más importante.
Muchos estudios genéticos han identificado mutaciones comunes, mutaciones raras y variaciones en el número de copias (VNC) en miles de niños con autismo; sin embargo, estos genes candidatos genéticos identificados suelen tener una baja prevalencia entre las poblaciones. Por ejemplo, sólo se han identificado variantes de CACNA1H en 6/~461 casos de autismo, y mutantes de CNTN4 en 7/~2000 casos [5].
Por otro lado, los factores ambientales pueden afectar directamente al desarrollo de grandes poblaciones de lactantes y niños pequeños a través de la neurotoxicidad y repercutir en la expresión génica modificando los estados epigenéticos, lo que influye enormemente en la prevalencia del autismo. Estudios a gran escala han descubierto que la exposición a factores ambientales nocivos, como las enfermedades maternas y el abuso de drogas, la exposición a la contaminación atmosférica, los comportamientos en la vida familiar y la exposición a metales durante el periodo intrauterino y las primeras etapas del desarrollo pueden desempeñar un papel clave en el desarrollo del TEA, además de otros factores (Figura 1).
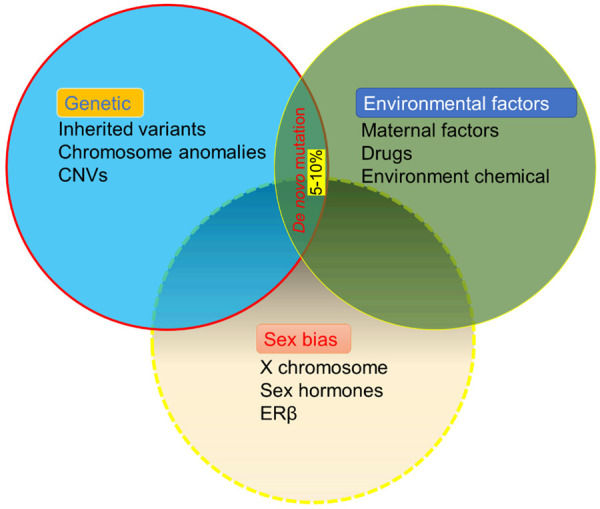
Figura 1. Las contribuciones de la genética, la exposición ambiental y los sesgos sexuales a la etiología de los trastornos del espectro autista. Se estima que los factores genéticos y ambientales contribuyen por igual al desarrollo de los TEA. Se han observado sesgos sexuales, con una prevalencia del autismo de 4 varones por cada mujer.
La diabetes materna induce autismo en la descendencia
Los trastornos metabólicos prenatales se asocian a una mayor prevalencia de TEA y otros trastornos en la descendencia. Un metaanálisis de Wan et al., por ejemplo, identificó pruebas de una asociación significativa entre la diabetes materna y los trastornos psiquiátricos en la descendencia [6]. Dado que la diabetes afecta hasta al 15% de las mujeres embarazadas en todo el mundo [7], debería prestarse mucha más atención al papel de la diabetes materna como factor de desarrollo del autismo en los niños y la descendencia.
Estudios epidemiológicos recientes han indicado que la diabetes materna, que incluye la diabetes tipo 1 (T1D), la diabetes tipo 2 (T2D) y la diabetes mellitus gestacional (DMG), diagnosticada antes de la 26ª semana de gestación, está muy asociada con un mayor riesgo de autismo en la descendencia, como se muestra en la Tabla 1. La diabetes tipo 2 preexistente en la madre también se asocia significativamente con el riesgo de TEA en la descendencia, mientras que el riesgo asociado es ligeramente inferior al riesgo de DMG a las 26 semanas [8-10].
Tabla 1. Contribución de los factores ambientales al desarrollo de los TEA (enlace a tabla en inglés)
Todos los análisis se realizaron utilizando el software estadístico SAS (SAS Institute Inc., Cary, NC, EE.UU.). La distribución de las características de los factores del embarazo se calculó en función de los estados autista y materno mediante pruebas t de muestras independientes de dos colas de las pruebas continuas y las pruebas chi-cuadrado de las variables categóricas. La asociación entre el estado materno y el TEA se determinó mediante un modelo de regresión logística multivariante para evaluar la odds ratio (OR) y el intervalo de confianza (IC) del 95%. N/D-No definido.
Sin embargo, el mecanismo detallado del modo en que la diabetes materna influye en el desarrollo de los TEA sigue sin estar claro. Nuestra investigación anterior, que se centró en el uso de modelos de diabetes en roedores, descubrió que las crías de madres diabéticas mostraban comportamientos similares al autismo, como una reducción de las vocalizaciones ultrasónicas y un deterioro de la interacción social.
Que sepamos, es la primera vez que se realiza un estudio de este tipo. Además, descubrimos que la superóxido dismutasa (SOD2) en la amígdala de las crías de las ratas estaba inhibida como resultado de la metilación de histonas mediada por el estrés oxidativo (Figura 2). Así pues, sugerimos que la hiperglucemia induce la generación de especies reactivas del oxígeno (ROS) persistentes y que la inhibición de la SOD2 es un posible mecanismo del comportamiento autista inducido por el embarazo diabético materno [11]. También se supone que la diabetes materna afecta al neurodesarrollo de la descendencia a través de procesos que incluyen la disfunción inmunitaria, el aumento del estrés oxidativo y la supresión de SOD2 en las células madre hematopoyéticas (HSC) [12].

Figura 2. La diabetes durante el embarazo induce comportamientos similares al autismo en la descendencia. A. Comportamientos similares al TEA inducidos por la diabetes materna en la descendencia. La sobreexpresión de SOD2 o de imitadores de SOD puede revertir parcialmente el comportamiento autista en la descendencia, pero la inhibición de la expresión de SOD2 inducirá el comportamiento autista. B. La hiperglucemia suprime la expresión de SOD2 a través de la metilación de histonas mediada por el estrés oxidativo. La metilación de histonas en H3K9me2 del promotor de SOD2 desencadena cambios epigenéticos, que posteriormente conducen a la inactivación de SOD2. La inhibición de la actividad de SOD2 agrava entonces la generación de ROS e induce estrés oxidativo.
Dilucidar el papel de la diabetes materna en la patogénesis de los TEA en la descendencia ayudará a replantearse la patogénesis del autismo desde la perspectiva de la interacción multifactorial, ampliando así la comprensión de la patogénesis de los TEA y proporcionando una diana para el diagnóstico y el tratamiento farmacológico de los TEA. Dado que la exposición excesiva a la glucosa en el útero puede provocar cambios fetales permanentes, la hiperglucemia materna puede tener efectos a largo plazo sobre el desarrollo y la función del cerebro fetal.
La obesidad y la diabetes maternas afectan al desarrollo del cerebro y el sistema nervioso del feto, lo que podría apuntar a un mecanismo de desarrollo del autismo. Pruebas anteriores han demostrado que la presencia de diabetes materna, además de obesidad y trastornos metabólicos relacionados, tanto antes como durante el embarazo, puede dañar el desarrollo normal del hipocampo y causar un desarrollo neuroconductual anormal en la descendencia [13].
Además, también hemos descubierto que la deficiencia de vitamina D empeora los fenotipos similares al autismo inducidos por la diabetes materna en la descendencia a través de la modificación epigenética causada por la diabetes materna [14]. Utilizando este modelo preestablecido de ratón autista inducido por diabetes materna, también hemos descubierto que las crías de madres con diabetes materna muestran una inhibición significativa del receptor de oxitocina (OXTR) en la región de la amígdala del tejido cerebral, mientras que los ratones knockout prenatales de OXTR muestran un empeoramiento de los síntomas de autismo inducido por diabetes materna.
Otros estudios han demostrado que la inhibición del OXTR está causada por un estrés oxidativo persistente y por la metilación debida a la hiperglucemia, que posteriormente provoca la disociación del receptor β de estrógenos (ERβ) del promotor del OXTR [15].
La exposición a sustancias químicas induce el autismo
La exposición a sustancias químicas es una de las condiciones del embarazo más comúnmente asociadas con efectos adversos en la descendencia. Por ejemplo, las mujeres embarazadas que consumieron marihuana durante el embarazo tenían 1,5 veces más probabilidades de dar a luz a un niño con autismo, y también se ha descubierto que el riesgo de autismo en la descendencia está asociado a la exposición materna a pesticidas [25-27]. La Tabla 1 enumera la exposición prenatal a sustancias químicas asociadas con el autismo. Sin embargo, se sabe poco sobre los mecanismos subyacentes de estas relaciones.
El uso de modelos animales puede servir para conocer mejor el comportamiento autista de la descendencia y ayudar a comprender los mecanismos del efecto de la exposición a xenobióticos durante el embarazo sobre el comportamiento autista. Se ha demostrado que la exposición materna a pesticidas modifica el desarrollo de las células neuronales y el comportamiento de las crías de animales. La investigación epidemiológica ha demostrado que la exposición materna al herbicida glifosato aumenta la incidencia del autismo en la descendencia; en uno de estos estudios, los investigadores expusieron a ratones preñados al glifosato y descubrieron que se producía un aumento de los comportamientos de tipo autista en la descendencia, incluidos déficits de interacción social y déficits cognitivos, además de una alteración de la composición de la microbiota intestinal.
Además, se observó un aumento significativo de la expresión de la epóxido hidrolasa soluble (sEH) en el cerebro de la descendencia tras la exposición materna al glifosato, mientras que los inhibidores de la sEH fueron capaces de restaurar los efectos de la exposición materna al glifosato en la inducción de comportamientos similares al autismo en la descendencia. Estos hallazgos indican que el aumento de sEH desempeña un papel vital en los comportamientos similares al TEA en la descendencia [28]. Así, el uso de modelos de rata o ratón para estudiar la exposición a factores ambientales durante el embarazo puede ayudar a simular los efectos adversos de la exposición a tóxicos ambientales que causan autismo en humanos.
Además, en lo que respecta a la exposición a metales, un estudio de Holmes et al. [29] realizó una prueba t de los niveles de plomo y mercurio en la sangre de 203 niños normales y descubrió que los niveles de plomo en sangre estaban positivamente correlacionados con el autismo, y los niveles de mercurio en sangre estaban relacionados con un aumento del comportamiento similar al autismo.
En otro estudio, se analizó la sangre de 34 y 35 niños con autismo, y se descubrió que los niveles de plomo y mercurio en los niños con autismo eran significativamente más altos que los niveles correspondientes en los controles. Los estudios han demostrado que los niveles de zinc y manganeso en el pelo están negativamente correlacionados con el autismo en los sujetos, y la gravedad de los síntomas del autismo está positivamente correlacionada con la presencia de ciertos metales pesados como el plomo y el mercurio [30-32].
Los desequilibrios hormonales maternos son un importante factor de riesgo de TEA
Los estudios epidemiológicos han demostrado que las anomalías hormonales en mujeres embarazadas son un importante factor de riesgo potencial de autismo en la descendencia y que las hormonas sexuales pueden ser parte de la causa del autismo. Realizamos un estudio epidemiológico de casos en la población de China y descubrimos que la progestina prenatal está estrechamente relacionada con la prevalencia de TEA. Esto incluye el uso de progesterona para prevenir el aborto, el uso de píldoras de progesterona y la ingesta materna de marisco contaminado con progestina.
Se ha demostrado que la progesterona modula las respuestas neurogénicas y perjudica el desarrollo de respuestas cognitivas al regular a la baja la expresión del ERβ. Se ha informado de que las expresiones del receptor de estrógeno β (ERβ) y de los cofactores del receptor de estrógeno estaban significativamente suprimidas en los cerebros de pacientes autistas [33].
Por lo tanto, se especula que la exposición prenatal a la progesterona sintética puede inducir comportamientos similares al autismo mediante la inhibición del ERβ en la descendencia. Un modelo in vivo de rata expuesta a progesterona mostró que la exposición prenatal a levonorgestrel inducía comportamientos autistas en la descendencia y suprimía la expresión de ERβ en el cerebro. Además, algunas investigaciones in vivo han revelado que los ratones ERβ knockout mostraban síntomas evidentes de ansiedad, déficits cognitivos y comportamiento depresivo [34].
Dado que se ha descubierto que la inhibición de ERβ induce el comportamiento autista, los agonistas del receptor de estrógeno β y/o la sobreexpresión de ERβ podrían ayudar a mejorar el comportamiento autista. Otras investigaciones descubrieron que los comportamientos similares al autismo inducidos por la exposición prenatal a la progesterona se rescataban mediante el tratamiento con resveratrol, un fármaco que activa el ERβ [8].
Además, se ha descubierto que un diagnóstico materno de síndrome de ovario poliquístico (SOP) aumenta el riesgo de TEA en la descendencia en un 59%, y se ha descubierto que el hiperandrogenismo prenatal induce comportamientos similares al autismo, como el deterioro del reconocimiento heterosexual y la disminución de la frecuencia de vocalización ultrasónica, en la descendencia [35].
Creemos que esto se debe a que las hormonas afectan al desarrollo de los sistemas neuroendocrino y neuroinmunitario en las primeras etapas de la vida, lo que conduce al desarrollo del TEA. Cada vez hay más pruebas que indican que la placenta puede ser especialmente importante como mediadora de las acciones de las hormonas endocrinas ambientales sobre el cerebro en desarrollo, lo que hace que la inhibición del ERβ inducida por hormonas en el cerebro masculino sea más sensible (Figura 3).
Comprender cómo los diversos factores de riesgo afectan al neurodesarrollo puede ayudar en el proceso de identificar la etiología de los TEA con mayor claridad. Nuestra investigación actual sugiere que la exposición prenatal a la dihidrotestosterona induce el desarrollo del TEA a través de la hipermetilación en el promotor del ERβ, que suprime la expresión del ERβ [36].
Anteriormente, hemos determinado que la progestina y la noretindrona pueden aumentar la dimetilación de la histona H3 lisina9 (H3K9me2) y la 3-metilación de H3K27 (H3K27me3) en el promotor ERβ en las neuronas de la amígdala, que es responsable del comportamiento similar al autismo inducido por la hormona materna [37].

Figura 3. La exposición materna a hormonas durante el embarazo está relacionada con un mayor riesgo de desarrollo de TEA en la descendencia. La exposición a dihidrotestosterona, progestágenos y noretindrona puede inducir una metilación significativa del promotor de ERβ e inhibir la expresión de ERβ en la descendencia.
El sesgo sexual como factor de riesgo significativo para el autismo
Hombres y mujeres tienen tendencias significativamente diferentes en el desarrollo de enfermedades neurológicas. Las mujeres son más propensas a trastornos del estado de ánimo como la ansiedad, la depresión y el trastorno bipolar, mientras que los hombres son más propensos a padecer trastornos como la enfermedad de Parkinson (EP), el trastorno por déficit de atención con hiperactividad (TDAH) y el autismo.
Un fenómeno interesante en la investigación del autismo es que el TEA afecta a los varones en mucha mayor proporción que a las mujeres. La prevalencia del autismo en los varones es aproximadamente cuatro veces superior a la de las mujeres [38], especialmente en los casos de autismo grave, en los que la proporción entre varones y mujeres es de casi 11 a 1. Además de estas diferencias en la prevalencia, los varones y las mujeres con autismo también tienden a mostrar síntomas diferentes. Los varones con autismo tienden a mostrar síntomas externos, como agresividad e hiperactividad, mientras que las mujeres con autismo tienden a mostrar menos comunicación social, además de comportamientos restrictivos y repetitivos [39].
Sin embargo, pocos investigadores prestan atención al sexo biológico en la búsqueda de factores que afectan al autismo. Una mejor comprensión de los mecanismos moleculares por los que las diferencias de sexo afectan al desarrollo de los cerebros autistas ayudará a diseñar las mejores estrategias de diagnóstico y tratamiento para cada sexo.
El profesor Donald W. Pfaff propone una «teoría de los tres golpes del autismo», es decir, que las interacciones entre genes, entorno y sexo sirven para inducir el desarrollo del TEA. Basándose en estudios realizados con el modelo de ratón Cntnap2 de autismo, los tres hits tienen diversos efectos sinérgicos sobre el comportamiento social y el reconocimiento social [40].
Un posible mecanismo de las diferencias de género en la prevalencia del autismo y la expresión de los síntomas puede entenderse a través de los diferentes efectos de las hormonas sexuales. Muchas investigaciones refuerzan la noción de que el estrógeno tiene un cierto efecto neuroprotector, y un estudio particularmente interesante ha descubierto que la expresión del receptor β del estrógeno en niños con autismo es muy baja y no es suficiente para mediar el efecto protector del estrógeno. Así, se sugiere que la expresión reducida del receptor beta de estrógenos, además de la inactivación de la enzima aromatasa que ayuda a convertir la testosterona en estrógeno, puede tener un impacto significativo en los individuos autistas y puede conducir a las mayores tasas de prevalencia del autismo observadas en los varones [33].
Además, algunos estudios interesantes han demostrado que los niveles de expresión proteica y génica del receptor huérfano alfa relacionado con el ácido retinoico (RORA) y la aromatasa están regulados a la baja en la corteza cerebral de los varones con autismo, y se especula que las deficiencias de RORA y aromatasa conducen a niveles más altos de testosterona, lo que puede aumentar el riesgo de TEA [41].
Así pues, la testosterona puede servir como factor específico masculino que aumente la susceptibilidad al autismo. En un modelo de pez cebra con mutantes del gen de riesgo de autismo CNTNAP2, se descubrió que los agonistas de los receptores de estrógenos pueden revertir la mutación del fenotipo conductual mutante, lo que indica que las hormonas sexuales contribuyen a los antecedentes genéticos [42].
Mediante un análisis de enriquecimiento de novo basado en el sexo, podemos identificar mutaciones genéticas candidatas que afectan a las diferencias entre hombres y mujeres. Recientemente, Eichler et al. investigaron cómo los genes del autismo difieren según el sexo, peinando el enriquecimiento de novo de 2.133 niñas y 4.641 niños diagnosticados de autismo. Descubrieron que sólo 17 genes eran exclusivos de las pacientes femeninas con autismo, y sólo 18 genes se encontraban exclusivamente en los varones. Otros 19 genes se encontraron en ambos sexos [43].
Por lo tanto, si hay genes en el cromosoma Y con enriquecimiento de novo, pueden servir para proporcionar información central para descubrir los factores sexuales que afectan al sesgo de género para el autismo. En este estudio, los autores también descubrieron que las mutaciones genéticas raras asociadas al TEA abundan en los casos femeninos en una proporción muy superior a la de los casos masculinos.
Se identificó así un efecto protector femenino (EPF) frente al comportamiento autista, y se predice que las mujeres pueden tener mutaciones de mayor gravedad o número sin que se desarrolle el TEA y pueden tolerar más variantes genéticas de riesgo para el autismo que los varones (Figura 4) [44].

Figura 4. Se especula que las hormonas sexuales y las variaciones genéticas desempeñan un papel importante en la patología del TEA tanto en hombres como en mujeres. La teoría de los tres golpes del autismo: genes, entorno y sexo, además del efecto protector femenino (EPF), pueden ser factores importantes que determinen las diferencias de género en la prevalencia del autismo y la expresión de los síntomas.
Conclusión y perspectivas de futuro
El TEA es un trastorno del neurodesarrollo que dura toda la vida, es irreversible e incurable y supone una gran carga económica, además de tener un impacto significativo en las familias y la sociedad. En la actualidad, las causas del TEA aún no se han esclarecido por completo, aunque se sabe que tanto los factores genéticos como los ambientales están implicados.
Según nuestras investigaciones y conocimientos, la mayoría de las investigaciones sobre el autismo realizadas hasta la fecha se centran principalmente en las variaciones genéticas, aunque muchos estudios epidemiológicos han establecido una fuerte asociación entre los factores ambientales prenatales y los resultados adversos del autismo en la descendencia.
Se sabe poco sobre los factores ambientales en la etiología del TEA. La influencia de la genética y el entorno es inseparable cuando se habla de los TEA, pero hay relativamente pocos estudios sobre los vínculos entre genes y entorno que afectan a los TEA.
Por lo tanto, se necesitan estudios más convincentes que respalden la influencia de la exposición ambiental simulada actual a la hora de provocar cambios epigenéticos que puedan conducir al desarrollo de los TEA. Creemos que los factores ambientales pueden gobernar la expresión génica cambiando marcadores epigenéticos como la metilación del ADN y la acetilación de histonas, siendo probablemente la metilación de histonas el marcador más importante, y afectando a las diferentes etapas de la vía biológica del desarrollo cerebral, incluyendo la diferenciación neuronal, la migración y la formación de tubos neurales, sinapsis y mielina, afectando finalmente a las funciones cognitivas y comunicativas de los individuos y desencadenando el TEA.
Analizar el desarrollo temprano desde la perspectiva de la epigenética nos permite mostrar cómo los factores ambientales pueden influir en el desarrollo del autismo en los niños, y enfoques como la secuenciación del ADN de nueva generación y los análisis epigenéticos pueden revelar aún más el mecanismo detallado de los factores ambientales que afectan a la transcripción de genes y las interacciones epigenéticas. Además, el uso de modelos neuronales derivados de células madre pluripotentes inducidas (iPSC) y/o la tecnología de organoides cerebrales para comprender los efectos de los factores ambientales en los procesos tempranos durante la neurogénesis aportará pistas sobre los orígenes del autismo (Figura 5).

Figura 5. Las iPSC o los organoides cerebrales proporcionan un sistema modelo para comprender la fisiopatología y los mecanismos patogenéticos subyacentes de los factores de riesgo maternos que pueden inducir autismo en la descendencia.
En última instancia, el objetivo de toda investigación sobre el autismo debe ser proporcionar estrategias de prevención e intervención con posibilidades de aplicación en el mundo real. Las futuras investigaciones encaminadas a comprender el papel de los factores ambientales en la etiología del TEA promoverán innovaciones que ayuden a alcanzar este objetivo.
Agradecimientos
This work was funded by the Science and Technology Innovation Committee of Shenzhen (No. JCYJ20200109150700942, No. JCYJ20180306170922163), the Key Realm R&D Program of Guangdong Province (2019B030335001), the Natural Science Foundation of Guangdong Province (2020A1515011581, 2021A1515010985), the Sanming Project of Medicine (No. SZSM201612079), the Shenzhen Fund for Guangdong Provincial High Level Clinical Key Specialties (No. SZGSP013), and the Shenzhen Key Medical Discipline Construction Fund (No. SZXK042).
Declaración de conflicto de intereses
Ninguno.
Referencias
1. Hyman SL, Levy SE, Myers SM. Identification, evaluation, and management of children with autism spectrum disorder. Pediatrics. 2020;145:e20193447. [PubMed] [Google Scholar]
2. Baio J, Wiggins L, Christensen DL, Maenner MJ, Daniels J, Warren Z, Kurzius-Spencer M, Zahorodny W, Robinson Rosenberg C, White T, Durkin MS, Imm P, Nikolaou L, Yeargin-Allsopp M, Lee LC, Harrington R, Lopez M, Fitzgerald RT, Hewitt A, Pettygrove S, Constantino JN, Vehorn A, Shenouda J, Hall-Lande J, Van Naarden Braun K, Dowling NF. Prevalence of autism spectrum disorder among children aged 8 years - autism and developmental disabilities monitoring network, 11 sites, United States, 2014. MMWR Surveill Summ. 2018;67:1–23. [PMC free article] [PubMed] [Google Scholar]
3. Ramaswami G, Geschwind DH. Genetics of autism spectrum disorder. Handb Clin Neurol. 2018;147:321–329. [PubMed] [Google Scholar]
4. Wassink TH, Piven J. The molecular genetics of autism. Curr Psychiatry Rep. 2000;2:170–175. [PubMed] [Google Scholar]
5. Ladd-Acosta C, Hansen KD, Briem E, Fallin MD, Kaufmann WE, Feinberg AP. Common DNA methylation alterations in multiple brain regions in autism. Mol Psychiatry. 2014;19:862–871. [PMC free article] [PubMed] [Google Scholar]
6. Wan H, Zhang C, Li H, Luan S, Liu C. Association of maternal diabetes with autism spectrum disorders in offspring: a systemic review and meta-analysis. Medicine (Baltimore) 2018;97:e9438. [PMC free article] [PubMed] [Google Scholar]
7. International Diabetes Federation. IDF policy briefing: diabetes in pregnancy: protecting maternal health. 2013 [Google Scholar]
8. Xie W, Ge X, Li L, Yao A, Wang X, Li M, Gong X, Chu Z, Lu Z, Huang X, Jiao Y, Wang Y, Xiao M, Chen H, Xiang W, Yao P. Resveratrol ameliorates prenatal progestin exposure-induced autism-like behavior through ERβ activation. Mol Autism. 2018;9:43. [PMC free article] [PubMed] [Google Scholar]
9. Xiang AH, Wang X, Martinez MP, Walthall JC, Curry ES, Page K, Buchanan TA, Coleman KJ, Getahun D. Association of maternal diabetes with autism in offspring. JAMA. 2015;313:1425–1434. [PubMed] [Google Scholar]
10. Li M, Fallin MD, Riley A, Landa R, Walker SO, Silverstein M, Caruso D, Pearson C, Kiang S, Dahm JL, Hong X, Wang G, Wang MC, Zuckerman B, Wang X. The association of maternal obesity and diabetes with autism and other developmental disabilities. Pediatrics. 2016;137:e20152206. [PMC free article] [PubMed] [Google Scholar]
11. Wang X, Lu J, Xie W, Lu X, Liang Y, Li M, Wang Z, Huang X, Tang M, Pfaff DW, Tang YP, Yao P. Maternal diabetes induces autism-like behavior by hyperglycemia-mediated persistent oxidative stress and suppression of superoxide dismutase 2. Proc Natl Acad Sci U S A. 2019;116:23743–23752. [PMC free article] [PubMed] [Google Scholar]
12. Lu J, Xiao M, Guo X, Liang Y, Wang M, Xu J, Liu L, Wang Z, Zeng G, Liu K, Li L, Yao P. Maternal diabetes induces immune dysfunction in autistic offspring through oxidative stress in hematopoietic stem cells. Front Psychiatry. 2020;11:576367. [PMC free article] [PubMed] [Google Scholar]
13. Menting MD, van de Beek C, Mintjens S, Wever KE, Korosi A, Ozanne SE, Limpens J, Roseboom TJ, Hooijmans C, Painter RC. The link between maternal obesity and offspring neurobehavior: a systematic review of animal experiments. Neurosci Biobehav Rev. 2019;98:107–121. [PubMed] [Google Scholar]
14. Liang Y, Yu H, Ke X, Eyles D, Sun R, Wang Z, Huang S, Lin L, McGrath JJ, Lu J, Guo X, Yao P. Vitamin D deficiency worsens maternal diabetes induced neurodevelopmental disorder by potentiating hyperglycemia-mediated epigenetic changes. Ann N Y Acad Sci. 2021;1491:74–88. [PubMed] [Google Scholar]
15. Liu J, Liang Y, Jiang X, Xu J, Sun Y, Wang Z, Lin L, Niu Y, Song S, Zhang H, Xue Z, Lu J, Yao P. Maternal diabetes-induced suppression of oxytocin receptor contributes to social deficits in offspring. Front Neurosci. 2021;15:634781. [PMC free article] [PubMed] [Google Scholar]
16. Sanchez CE, Barry C, Sabhlok A, Russell K, Majors A, Kollins SH, Fuemmeler BF. Maternal pre-pregnancy obesity and child neurodevelopmental outcomes: a meta-analysis. Obes Rev. 2018;19:464–484. [PMC free article] [PubMed] [Google Scholar]
17. Cherskov A, Pohl A, Allison C, Zhang H, Payne RA, Baron-Cohen S. Polycystic ovary syndrome and autism: a test of the prenatal sex steroid theory. Transl Psychiatry. 2018;8:136. [PMC free article] [PubMed] [Google Scholar]
18. Kosidou K, Dalman C, Widman L, Arver S, Lee BK, Magnusson C, Gardner RM. Maternal polycystic ovary syndrome and the risk of autism spectrum disorders in the offspring: a population-based nationwide study in Sweden. Mol Psychiatry. 2016;21:1441–1448. [PMC free article] [PubMed] [Google Scholar]
19. Brown HK, Ray JG, Wilton AS, Lunsky Y, Gomes T, Vigod SN. Association between serotonergic antidepressant use during pregnancy and autism spectrum disorder in children. JAMA. 2017;317:1544–1552. [PubMed] [Google Scholar]
20. Maher GM, O’Keeffe GW, Kearney PM, Kenny LC, Dinan TG, Mattsson M, Khashan AS. Association of hypertensive disorders of pregnancy with risk of neurodevelopmental disorders in offspring: a systematic review and meta-analysis. JAMA Psychiatry. 2018;75:809–819. [PMC free article] [PubMed] [Google Scholar]
21. Jiang HY, Xu LL, Shao L, Xia RM, Yu ZH, Ling ZX, Yang F, Deng M, Ruan B. Maternal infection during pregnancy and risk of autism spectrum disorders: a systematic review and meta-analysis. Brain Behav Immun. 2016;58:165–172. [PubMed] [Google Scholar]
22. Brown AS, Cheslack-Postava K, Rantakokko P, Kiviranta H, Hinkka-Yli-Salomäki S, McKeague IW, Surcel HM, Sourander A. Association of maternal insecticide levels with autism in offspring from a national birth cohort. Am J Psychiatry. 2018;175:1094–1101. [PMC free article] [PubMed] [Google Scholar]
23. Philippat C, Barkoski J, Tancredi DJ, Elms B, Barr DB, Ozonoff S, Bennett DH, Hertz-Picciotto I. Prenatal exposure to organophosphate pesticides and risk of autism spectrum disorders and other non-typical development at 3 years in a high-risk cohort. Int J Hyg Environ Health. 2018;221:548–555. [PMC free article] [PubMed] [Google Scholar]
24. Fountain C, Zhang Y, Kissin DM, Schieve LA, Jamieson DJ, Rice C, Bearman P. Association between assisted reproductive technology conception and autism in California, 1997-2007. Am J Public Health. 2015;105:963–971. [PMC free article] [PubMed] [Google Scholar]
25. Corsi DJ, Donelle J, Sucha E, Hawken S, Hsu H, El-Chaâr D, Bisnaire L, Fell D, Wen SW, Walker M. Maternal cannabis use in pregnancy and child neurodevelopmental outcomes. Nat Med. 2020;26:1536–1540. [PubMed] [Google Scholar]
26. Wood AG, Nadebaum C, Anderson V, Reutens D, Barton S, O’Brien TJ, Vajda F. Prospective assessment of autism traits in children exposed to antiepileptic drugs during pregnancy. Epilepsia. 2015;56:1047–1055. [PubMed] [Google Scholar]
27. von Ehrenstein OS, Ling C, Cui X, Cockburn M, Park AS, Yu F, Wu J, Ritz B. Prenatal and infant exposure to ambient pesticides and autism spectrum disorder in children: population based case-control study. BMJ. 2019;364:l962. [PMC free article] [PubMed] [Google Scholar]
28. Pu Y, Yang J, Chang L, Qu Y, Wang S, Zhang K, Xiong Z, Zhang J, Tan Y, Wang X, Fujita Y, Ishima T, Wang D, Hwang SH, Hammock BD. Maternal glyphosate exposure causes autism-like behaviors in offspring through increased expression of soluble epoxide hydrolase. Proc Natl Acad Sci U S A. 2020;117:11753–11759. [PMC free article] [PubMed] [Google Scholar]
29. Holmes AS, Blaxill MF, Haley BE. Reduced levels of mercury in first baby haircuts of autistic children. Int J Toxicol. 2003;22:277–285. [PubMed] [Google Scholar]
30. Majewska MD, Urbanowicz E, Rok-Bujko P, Namyslowska I, Mierzejewski P. Age-dependent lower or higher levels of hair mercury in autistic children than in healthy controls. Acta Neurobiol Exp (Wars) 2010;70:196–208. [PubMed] [Google Scholar]
31. Lakshmi Priya MD, Geetha A. Level of trace elements (copper, zinc, magnesium and selenium) and toxic elements (lead and mercury) in the hair and nail of children with autism. Biol Trace Elem Res. 2011;142:148–158. [PubMed] [Google Scholar]
32. Geier DA, Kern JK, King PG, Sykes LK, Geier MR. Hair toxic metal concentrations and autism spectrum disorder severity in young children. Int J Environ Res Public Health. 2012;9:4486–4497. [PMC free article] [PubMed] [Google Scholar]
33. Crider A, Thakkar R, Ahmed AO, Pillai A. Dysregulation of estrogen receptor beta (ERβ), aromatase (CYP19A1), and ER co-activators in the middle frontal gyrus of autism spectrum disorder subjects. Mol Autism. 2014;5:46. [PMC free article] [PubMed] [Google Scholar]
34. Krezel W, Dupont S, Krust A, Chambon P, Chapman PF. Increased anxiety and synaptic plasticity in estrogen receptor beta-deficient mice. Proc Natl Acad Sci U S A. 2001;98:12278–12282. [PMC free article] [PubMed] [Google Scholar]
35. Xu XJ, Zhang HF, Shou XJ, Li J, Jing WL, Zhou Y, Qian Y, Han SP, Zhang R, Han JS. Prenatal hyperandrogenic environment induced autistic-like behavior in rat offspring. Physiol Behav. 2015;138:13–20. [PubMed] [Google Scholar]
36. Xiang D, Lu J, Wei C, Cai X, Wang Y, Liang Y, Xu M, Wang Z, Liu M, Wang M, Liang X, Li L, Yao P. Berberine ameliorates prenatal dihydrotestosterone exposure-induced autism-like behavior by suppression of androgen receptor. Front Cell Neurosci. 2020;14:87. [PMC free article] [PubMed] [Google Scholar]
37. Zou Y, Lu Q, Zheng D, Chu Z, Liu Z, Chen H, Ruan Q, Ge X, Zhang Z, Wang X, Lou W, Huang Y, Wang Y, Huang X, Liu Z, Xie W, Zhou Y, Yao P. Prenatal levonorgestrel exposure induces autism-like behavior in offspring through ERβ suppression in the amygdala. Mol Autism. 2017;8:46. [PMC free article] [PubMed] [Google Scholar]
38. Halladay AK, Bishop S, Constantino JN, Daniels AM, Koenig K, Palmer K, Messinger D, Pelphrey K, Sanders SJ, Singer AT, Taylor JL, Szatmari P. Sex and gender differences in autism spectrum disorder: summarizing evidence gaps and identifying emerging areas of priority. Mol Autism. 2015;6:36. [PMC free article] [PubMed] [Google Scholar]
39. Frazier TW, Georgiades S, Bishop SL, Hardan AY. Behavioral and cognitive characteristics of females and males with autism in the Simons Simplex Collection. J Am Acad Child Adolesc Psychiatry. 2014;53:329–340. e1–3. [PMC free article] [PubMed] [Google Scholar]
40. Schaafsma SM, Gagnidze K, Reyes A, Norstedt N, Månsson K, Francis K, Pfaff DW. Sex-specific gene-environment interactions underlying ASD-like behaviors. Proc Natl Acad Sci U S A. 2017;114:1383–1388. [PMC free article] [PubMed] [Google Scholar]
41. Hu VW, Sarachana T, Sherrard RM, Kocher KM. Investigation of sex differences in the expression of RORA and its transcriptional targets in the brain as a potential contributor to the sex bias in autism. Mol Autism. 2015;6:7. [PMC free article] [PubMed] [Google Scholar]
42. Hoffman EJ, Turner KJ, Fernandez JM, Cifuentes D, Ghosh M, Ijaz S, Jain RA, Kubo F, Bill BR, Baier H, Granato M, Barresi MJF, Wilson SW, Rihel J, State MW, Giraldez AJ. Estrogens suppress a behavioral phenotype in zebrafish mutants of the autism risk gene, CNTNAP2. Neuron. 2016;89:725–733. [PMC free article] [PubMed] [Google Scholar]
43. Turner TN, Wilfert AB, Bakken TE, Bernier RA, Pepper MR, Zhang Z, Torene RI, Retterer K, Eichler EE. Sex-based analysis of de novo variants in neurodevelopmental disorders. Am J Hum Genet. 2019;105:1274–1285. [PMC free article] [PubMed] [Google Scholar]
44. Robinson EB, Lichtenstein P, Anckarsäter H, Happé F, Ronald A. Examining and interpreting the female protective effect against autistic behavior. Proc Natl Acad Sci U S A. 2013;110:5258–5262. [PMC free article] [PubMed] [Google Scholar]
Artículo original en inglés:



Comentarios